2025. 8. 5. 오전 9:43:00
‘성가신 친구’는 ‘흡연’만큼 당신을 늙게 한다
출처: 한겨레 ( 한국 / 한국어 )
DNA 메틸화는 DNA 염기에 메틸기가 부착되는 화학 반응으로, 진핵생물에서 유전자 발현 조절, 세포 분화, 발생, 질병 등 다양한 생물학적 과정에 관여한다. 포유류에서는 주로 CpG 부위의 시토신에 메틸화가 일어나며, 식물에서는 CpG, CpHpG, CpHpH 부위에서 시토신이 메틸화될 수 있다. DNA 메틸화는 DNA 메틸전이효소(DNMT)에 의해 촉매되며, DNMT1은 메틸화 패턴 유지, DNMT3A와 DNMT3B는 새로운 메틸화 패턴 확립에 관여한다. DNA 메틸화는 유전자 발현 조절, 유전체 각인, X 염색체 불활성화, 전이인자 억제 등 다양한 기능을 수행하며, 암, 동맥경화증, 심부전, 노화, 신경 발달 질환 등 여러 질병과 관련이 있다. DNA 메틸화 연구 방법으로는 질량 분석법, 메틸화 특이적 PCR, 비스울파이트 시퀀싱 등이 사용되며, 곤충, 균류, 세균 등 다양한 생물에서도 DNA 메틸화가 발견된다.
| DNA 메틸화 | |
|---|---|
| 지도 정보 | |
| 기본 정보 | |
| 유형 | 생물학적 과정 |
| 관련 생물학적 과정 | 유전체 조절 |
| 특징 | |
| 설명 | DNA에서 시토신 또는 아데닌 염기의 화학적 변형으로, 유전체 조절에 중요한 역할. |
| 시토신 메틸화 | 포유류에서 가장 일반적인 형태, 주로 CpG 부위에서 발생. |
| 아데닌 메틸화 | 일부 박테리아와 진핵생물에서 발견, 최근 포유류에서도 보고됨. |
| 효소 | DNA 메틸트랜스퍼라제 (DNMT)에 의해 촉매됨. |
| 효과 | 유전자 발현 조절, 염색체 안정성 유지, 세포 분화 및 발생 조절 등 다양한 생물학적 과정에 영향. |
| 연구 | 후성유전체 연구의 중요한 부분, 암 및 기타 질병 연구에 활용. |
| 시토신 메틸화 상세 정보 | |
| 위치 | 주로 시토신 염기의 5번 탄소 (5mC) CpG 디뉴클레오타이드에서 자주 발생 |
| 기능 | 유전자 발현 억제 염색체 구조 조절 DNA 복제 및 복구 과정에 관여 |
| 패턴 | 세포 유형 특이적 발생 단계에 따라 변화 |
| 연구 방법 | 바이설파이트 시퀀싱 메틸화 특이적 PCR (MSP) 면역침강 |
| 관련 질병 | 암 발달 장애 신경 질환 |
| 아데닌 메틸화 상세 정보 | |
| 위치 | 아데닌 염기의 N6 위치 (6mA) |
| 기능 | DNA 복제 전사 조절 DNA 손상 복구 |
| 연구 | 액체 크로마토그래피-질량 분석법 (LC-MS/MS) |
| 종별 메틸화 특징 | |
| 포유류 | 시토신 메틸화 (5mC)가 주요 형태 CpG 부위에서 주로 발견 배아 발생, 조직 특이적 발현에 중요 |
| 식물 | 시토신 메틸화 (5mC)가 일반적 CpG, CHG, CHH 부위에서 발견 (H는 A, C 또는 T) |
| 곰팡이 | 일부 종에서 시토신 메틸화 발견 예를 들어, Neurospora crassa는 RIP (Repeat-Induced Point Mutation) 과정에서 소량의 메틸화된 DNA를 가짐 Dictyostelium discoideum은 DNA 메틸화가 거의 없음 |
| 효모 | Saccharomyces cerevisiae와 Schizosaccharomyces pombe는 DNA 메틸화가 없음 |
| 초파리 | Drosophila melanogaster에서 시토신 메틸화 발견 |
| 선충 | Caenorhabditis elegans에서 메틸화된 시토신 및 하이드록시메틸시토신이 발견됨 |
| 추가 정보 | |
| 역사적 연구 | Physarum polycephalum에서 초기 DNA 메틸화 연구 진행 |
DNA 메틸화는 1948년 롤린 호치키스(Rollin Hotchkiss)에 의해 처음 발견되었다.[136] 1970년대에는 제한 효소와 DNA 메틸화의 관계가 밝혀졌다. 1980년대에는 DNA 메틸화가 유전자 발현 조절에 중요한 역할을 한다는 사실이 밝혀졌다. 1990년대 이후 DNA 메틸화와 관련된 다양한 연구가 진행되고 있으며, 암을 비롯한 여러 질병과의 연관성이 밝혀지고 있다.[137]
DNA 메틸화는 DNA 염기 중 시토신(C) 또는 아데닌(A)에 메틸기(-CH3)가 붙는 화학 반응이다. 포유류에서는 주로 CpG 부위(시토신과 구아닌이 인산기로 연결된 부위)의 시토신에 메틸화가 일어난다. 식물에서는 CpG, CpHpG, CpHpH 부위의 시토신에 메틸화가 일어난다 (H는 A, C, T 중 하나).[76] DNA 메틸화는 DNA 메틸전이효소(DNMT)에 의해 촉매된다.
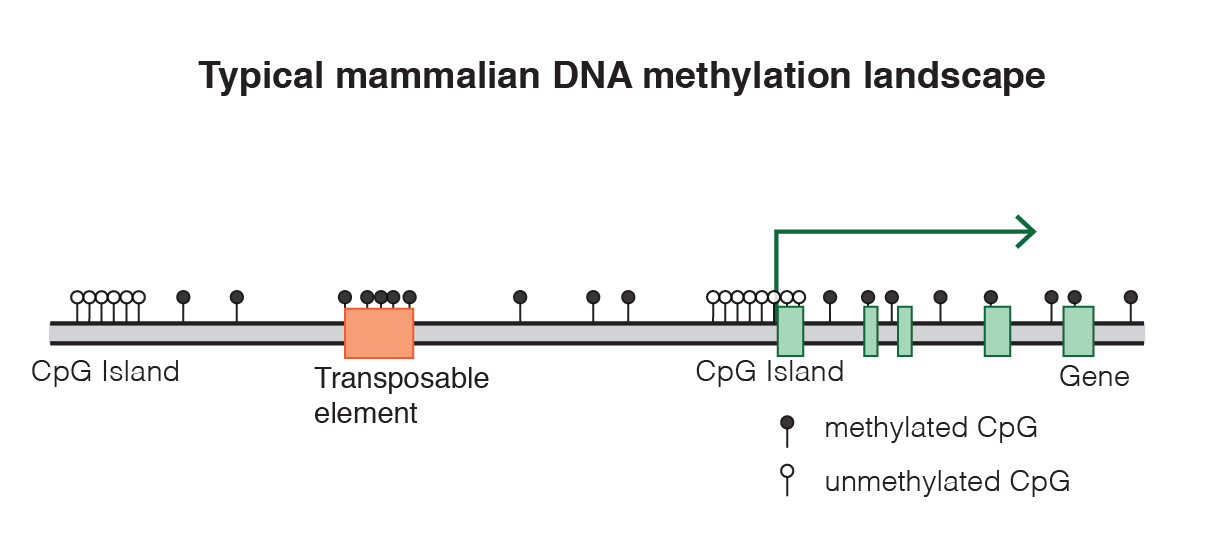
척추동물의 DNA 메틸화 지형은 다른 생물에 비해 매우 특이하다. 포유류의 경우, 체세포 CpG 이중 핵산염기의 약 75%가 메틸화되어 있으며, DNA 메틸화는 특정 위치에서 특이적으로 제외되어야 하는 기본 상태로 나타난다.[19] [16][20] 반면, 대부분의 식물, 무척추동물, 균류 또는 원생생물의 게놈은 메틸화된 영역과 메틸화되지 않은 영역이 번갈아 나타나는 "모자이크" 메틸화 패턴을 보이며, 특정 게놈 요소만을 표적으로 한다.[21][22]
3. 생화학적 특징
메틸화된 시토신(C) 잔기는 시간이 지남에 따라 자발적으로 탈아미노화되어 티민(T) 잔기를 형성한다. 따라서 CpG 이중 핵산염기는 꾸준히 TpG 이중 핵산염기로 탈아미노화되며, 이는 인간 게놈에서 CpG 이중 핵산염기의 과소 표현(예상 빈도의 21%만 존재)으로 나타난다.[23] 반면, 메틸화되지 않은 C 잔기의 자발적 탈아미노화는 우라실(U) 잔기를 생성하며, 이는 세포에 의해 신속하게 인식되고 복구된다.
4. 생물학적 기능
포유류 게놈에서 높은 CpG 메틸화는 자발적 돌연변이 빈도를 증가시키기 때문에 진화적 비용이 따른다. 시토신의 아미노기 손실은 높은 빈도로 발생하며, 메틸화 여부에 따라 결과가 다르다. 메틸화된 C 잔기는 시간이 지남에 따라 자발적으로 탈아미노화되어 T 잔기를 형성한다. 따라서 CpG 이중 핵산염기는 꾸준히 TpG 이중 핵산염기로 탈아미노화되며, 이는 인간 게놈에서 CpG 이중 핵산염기의 과소 표현(예상 빈도의 21%만 존재)으로 입증된다.[23] 반면, 메틸화되지 않은 C 잔기의 자발적 탈아미노화는 U 잔기를 생성하며, 이는 세포에 의해 신속하게 인식되고 복구된다.
DNA 메틸화는 시토신(Cytosine)의 피리미딘(Pyrimidine) 고리 5번 탄소 원자 또는 아데닌(Adenine)의 퓨린(Purine) 고리 6번 질소 원자에 메틸기(Methyl group)가 첨가되는 반응이다(시토신과 아데닌은 DNA를 구성하는 4가지 염기 중 2가지이다). 이러한 변형은 세포 분열을 거쳐도 유전된다. 일반적으로 DNA 메틸화는 접합자 형성 과정에서 제거되고 발생 과정에서의 이후 세포 분열을 통해 재구축된다. 그러나 최근 연구에 따르면 접합자에서는 메틸기의 완전한 제거보다는 메틸기의 수산화(Hydroxylation)가 일어나는 것으로 나타났다.[136]
DNA 메틸화는 또한 거의 모든 종류의 암 발생에 매우 중요한 역할을 한다.[137] DNA 메틸화는 정상적인 발생에 필수적이며, 각인 유전 및 X 염색체 불활성화, 반복 서열의 억제, 발암 등 많은 중요 단계와 관련이 있다.
비메틸화 CpG는 종종 많은 유전자의 5' 조절 영역에 존재하는 '''CpG 섬'''이라고 불리는 클러스터로 모여 있다. 암 등 많은 질병 과정에서 유전자 프로모터인 CpG 섬이 비정상적인 과메틸화를 받아 결과적으로 세포 분열에 의한 자세포에 전달되는 유전자 침묵이 일어난다. DNA 메틸화의 변화는 암 발생의 중요한 요소로 인식되고 있다. 과메틸화가 프로모터와 관련되어 유전자(종양 억제 유전자) 침묵을 일으키는 반면, 저메틸화는 일반적으로 초기 발생하는 염색체 불안정성이나 각인 상실과 관련이 있다. 그러나 저메틸화는 후성유전학적 치료의 표적이 될 수도 있다.[144]
지금까지의 연구에 따르면, 인간의 장기 기억 유지가 DNA 메틸화에 의해 조절된다는 것이 시사되고 있다.[145][146]
4. 1. 유전자 발현 조절
DNA 메틸화는 DNA에 메틸기가 첨가되는 반응으로, 주로 시토신의 5번 탄소 위치에 메틸기가 붙는 경우 유전자 발현이 감소하는 효과가 있다. DNA 메틸화는 초기 진핵생물 조상에서부터 존재했을 가능성이 높으며, 분석된 거의 모든 생물체에서 프로모터 영역의 메틸화는 유전자 발현 억제와 관련이 있다.[21][28] 활발하게 전사되는 유전자의 CpG가 풍부한 프로모터는 메틸화되지 않지만, 전사가 억제된 유전자가 반드시 메틸화된 프로모터를 가지는 것은 아니다.
DNA 메틸화는 두 가지 방식으로 유전자 전사에 영향을 줄 수 있다.[32]
특히, MeCP2의 손실은 레트 증후군과 관련이 있으며, MBD2는 암에서 과메틸화된 유전자의 전사 침묵을 매개한다.
DNA 메틸화는 전이인자의 영구적인 침묵을 보장하는 데 중요한 역할을 한다.[33] 전이인자 조절은 DNA 메틸화의 가장 오래된 기능 중 하나로 여겨지며, DNA 메틸화가 이 목적으로 진화했다는 주장도 있다.[34][35]
유전자 본체 메틸화는 스플라이싱 조절[37] 및 유전자 내 전사 단위 활성 억제[38]에 관여하는 것으로 보인다. 유전자 본체 메틸화는 H3K36 메틸화와 밀접하게 관련되어 있으며, 이는 크로마틴을 응축시켜 암호성 시작 부위의 활성화를 방지한다.[39]
4. 2. 세포 분화 및 발생
DNA 메틸화는 세포 분화 및 발생 과정에서 핵심적인 역할을 수행한다. 발생 초기 단계에서 DNA 메틸화 패턴이 재확립되며, 이는 세포의 운명을 결정하는 데 중요한 역할을 한다.[40] 특정 유전자의 메틸화 패턴은 세포 유형에 따라 다르게 나타나며, 이는 조직 특이적인 유전자 발현을 조절한다.[40]
포유류에서는 세대 간에 DNA 메틸화 패턴이 대부분 지워졌다가 다시 확립된다. 부모로부터 물려받은 거의 모든 메틸화는 생식세포형성 과정과 초기 배아발생 과정에서 탈메틸화와 재메틸화를 통해 지워진다.[40] 초기 배아 발생에서 탈메틸화는 수정란, 상실배, 포배에서 일어나며, 이후 착상 단계에서 메틸화가 일어나 CpG 섬은 메틸화로부터 보호된다. 이는 하우스키핑 유전자가 모든 세포에서 발현되도록 한다.[40] 착상 후 단계에서는 메틸화 패턴이 단계 및 조직 특이적으로 변화하며, 각 세포 유형을 정의하는 변화는 오랜 기간 안정적으로 유지된다.[40] 쥐의 사지 돌기 연구에서는 발생 과정에서 DNA 메틸화의 역동적인 특성이 관찰되었으며, 이는 기관 형성 과정에서 메틸화의 복잡한 조절과 재생 의학에 대한 잠재적 의미를 강조한다.[41]
DNA 메틸화는 유전체 각인과 X 염색체 불활성화에서 단일 대립 유전자 침묵 유지에 중요하며,[42][43] DNA 메틸화 손실은 각인 손실과 체세포에서 Xist의 재발현을 초래한다. 배아 발생 과정에서 생식세포 특이적 발현 유전자를 제외한 소수의 유전자만이 메틸화 상태를 변화시킨다.[44] 분화된 세포에서 DNA 메틸화는 필수적이며, DNA 메틸전이효소 녹아웃은 배아 또는 분만 후 치사율을 유발한다. 반면, 미분화 세포 유형에서는 DNA 메틸화가 불필요하다.
유전체 각인 현상으로 인해 모계 및 부계 게놈은 차별적으로 표시되며, 생식세포를 통과할 때마다 DNA 메틸화 재프로그래밍을 통해 재프로그래밍되어야 한다. 수정 후에는 부계 및 모계 게놈이 탈메틸화 및 재메틸화되지만, 각인된 유전자와 관련된 차별적 메틸화 영역은 제외된다. 이러한 재프로그래밍은 배아의 전능성과 획득된 후생유전학적 변화의 지움에 필요하다.[45]
B 세포 분화 주기 연구에 따르면, 초기 단계부터 분화된 단계까지 저메틸화가 나타나며, 생식 중심 B 세포와 기억 B 세포 단계 사이에서 가장 큰 메틸화 차이가 나타난다. 또한 B 세포 종양과 장수명 B 세포는 DNA 메틸화 서명에서 유사성을 보인다.[18]
DNA 메틸화는 정상적인 발생과 세포 분화에 중요하며, 세포 기억을 통해 유전자 발현 패턴을 안정적으로 변화시키거나 유전자 발현을 감소시킨다.
4. 3. 유전체 각인
유전체 각인은 부모 중 어느 쪽에서 유래한 유전자인지에 따라 유전자 발현이 달라지는 현상이다. 생식세포형성 과정에서 원시 생식세포는 부모에게서 받은 DNA 메틸화 패턴을 지우고, 전달하는 부모의 성별에 따라 다시 설정한다. 수정 후, 부계와 모계 게놈은 다시 탈메틸화와 재메틸화 과정을 거친다. (단, 각인된 유전자와 관련된 차별적으로 메틸화된 영역은 제외). 이러한 재프로그래밍은 새로 형성된 배아의 전능성과 획득된 후생유전학적 변화를 제거하는데 필요하다.[45]
DNA 메틸화는 유전체 각인과 X 염색체 불활성화에서 단일 대립 유전자 침묵을 유지하는 데 중요하다.[42][43] 발현되는 대립 유전자와 침묵되는 대립 유전자는 메틸화 상태가 다르며, DNA 메틸화가 소실되면 각인이 소실되고 체세포에서 Xist의 재발현이 나타난다.
4. 4. X 염색체 불활성화
포유류 암컷은 X 염색체를 두 개 가지고 있는데, 이 중 하나가 불활성화되는 현상을 X 염색체 불활성화라고 한다. 이는 DNA 메틸화와 밀접하게 관련되어 있으며, 유전자 발현의 균형을 유지하는 데 중요한 역할을 한다.
DNA 메틸화는 정상적인 발생에 필수적이며, X 염색체 불활성화와 같은 여러 중요한 단계와 관련이 있다.[136]
4. 5. 전이인자 억제
DNA 메틸화는 전이인자의 영구적인 억제를 보장하는 데 매우 적합하다.[33] 전이인자 조절은 동물, 식물 및 여러 원생생물이 공유하는 DNA 메틸화의 가장 오래된 기능 중 하나이다.[34] DNA 메틸화가 바로 이 목적, 즉 전이인자 억제를 위해 진화했다는 주장도 있다.[35]
4. 6. 게놈 확장
DNA 메틸화는 시간이 지남에 따라 숙주의 게놈에 통합된 바이러스나 기타 유해 요소의 유전자 발현을 억제한다.[137] 특히 전이인자의 DNA 메틸화는 게놈 확장과 관련이 있는 것으로 알려져 있다.
DNA 메틸화는 정상적인 발생 과정에 필수적이며, 각인 유전, X 염색체 불활성화, 반복 서열 억제, 발암 등 여러 중요한 단계와 관련이 있다. 포유류에서는 CpG 부위의 60~90%가 메틸화되어 있으며,[141][142] 메틸화된 시토신은 자발적인 탈아미노화 반응으로 티민으로 변이될 수 있다. 비메틸화 CpG는 주로 유전자의 5' 조절 서열에 위치한 CpG 섬에 모여 있다.
DNA 메틸화는 암을 비롯한 여러 질병과 연관되어 있다. 암과 같은 많은 질병 과정에서 유전자 프로모터 CpG 섬의 비정상적인 과메틸화는 세포 분열을 통해 딸세포에 전달되는 유전자 사일렌싱을 유발한다.[46] DNA 메틸화의 변화는 암 발생의 중요한 요소로, 과메틸화는 프로모터와 관련되어 종양 억제 유전자 침묵을 일으키는 반면, 저메틸화는 일반적으로 초기 발생하는 염색체 불안정성이나 각인 상실과 관련이 있다.[47]
DNA 메틸화는 두 가지 방식으로 유전자 전사에 영향을 미친다. 첫째, DNA 메틸화 자체가 전사 인자의 결합을 물리적으로 방해할 수 있다. 둘째, 메틸화된 DNA는 메틸-CpG 결합 도메인 단백질(MBD)과 결합하고, MBD 단백질은 히스톤 탈아세틸화 효소 및 기타 크로마틴 재구성 단백질을 유전자좌에 모집하여 불활성화된 크로마틴을 형성한다.[144] 특히, MeCP2의 결손은 레트 증후군과 관련이 있으며, MBD2는 암에서 과메틸화 유전자의 전사 억제를 매개한다.
인간의 장기 기억 유지가 DNA 메틸화에 의해 조절된다는 연구 결과도 있다.[145][146]
포유류에서 DNA 메틸화를 촉매하는 효소는 DNA 메틸전이효소(DNMT)이다. 포유류에는 DNMT1, DNMT3A, DNMT3B, DNMT3L 등 여러 종류의 DNMT가 존재한다.[72]
DNMT1은 DNA 복제 과정에서 기존의 메틸화 패턴을 딸 가닥에 복사하여 유지하는 역할을 하는 유지 메틸전이효소로 제안되었다. DNMT1이 결핍된 쥐 모델은 배아 9일째에 치사하는데, 이는 포유류 세포 발달에 DNMT1 활성이 필수적임을 보여준다.
DNMT3A와 DNMT3B는 발생 초기에 새로운 메틸화 패턴을 확립하는 *de novo* 메틸전이효소로 생각된다. DNMT3L은 촉매 활성이 없는 단백질이지만, *de novo* 메틸전이효소와 결합하여 DNA 결합 능력을 높이고 활성을 돕는다.[72]
쥐와 생쥐는 Muroidea 설치류의 공통 조상에서 탠덤 중복(tandem duplication)에 의해 ''Dnmt3b''의 파라로그(paralog)로 진화한 DNMT3C라는 세 번째 기능적 *de novo* 메틸전이효소 효소를 가지고 있다. DNMT3C는 초기 정자 형성 중에 전좌 가능 요소(transposable element)의 프로모터 메틸화를 촉매하며, 이는 이들의 후생유전적 억제와 수컷 생식력에 필수적인 것으로 나타났다.[73][74]
DNMT2 (TRDMT1)는 DNA가 아닌 아스파르트산 tRNA의 안티코돈 루프에 존재하는 시토신-38을 메틸화한다.[75]
| 효소 | 기능 |
|---|---|
| DNMT1 | DNA 복제 과정에서 기존 메틸화 패턴을 유지 |
| DNMT3A | 발생 과정에서 새로운 메틸화 패턴 확립 |
| DNMT3B | 발생 과정에서 새로운 메틸화 패턴 확립 |
| DNMT3L | 촉매 활성은 없지만, DNMT3A와 DNMT3B의 활성을 도움 |
| DNMT2 (TRDMT1) | 아스파르트산 tRNA의 안티코돈 루프에 존재하는 시토신-38을 메틸화, DNA 메틸화와 관련 없음 |
DNA 메틸화는 현재 과학 연구에서 사용되는 다음과 같은 분석법으로 검출할 수 있다.[101]
전통적인 방법
DNA 메틸화는 시토신의 피리미딘 고리 5번 탄소 원자 또는 아데닌의 퓨린 고리 6번 질소 원자에 메틸기(Methyl group)가 첨가되는 반응이다. 이러한 변형은 세포 분열을 거쳐도 유전된다. 일반적으로 DNA 메틸화는 접합자 형성 과정에서 제거되고 발생 과정에서의 이후 세포 분열을 통해 재구축되지만, 최근 연구에 따르면 접합자에서는 메틸기의 완전한 제거보다는 메틸기의 수산화(Hydroxylation)가 일어나는 것으로 나타났다.[136]
DNA 메틸화는 세포가 "자신이 어디에 있는지"를 기억(세포 기억)할 수 있도록 안정적으로 유전자 발현 패턴을 변화시키거나 유전자 발현을 감소시킨다. 예를 들어, 이자의 랑게르한스섬이 되도록 프로그램된 세포는 지속적으로 신호를 받지 않더라도 랑게르한스섬으로 남는다. 또한 DNA 메틸화는 시간이 지남에 따라 숙주의 게놈(Genome)에 통합된 바이러스(Virus)나 기타 유해 요소의 유전자 발현을 억제하며, 크로마틴(Chromatin) 구조의 기반을 형성하여 단일 DNA 서열에서 다세포 생물에 필요한 수많은 특징을 형성할 수 있게 한다. DNA 메틸화는 거의 모든 종류의 암(Cancer) 발생에 매우 중요한 역할을 한다.[137]
DNA 메틸화는 DNA에 메틸기가 첨가되는 것을 수반하며, 시토신의 피리미딘 고리 5번 탄소 원자에 메틸기가 붙으면 유전자 발현 감소라는 특이적인 효과가 있다. 시토신 5번 위치의 메틸화는 조사된 모든 척추동물(Vertebrate)에서 발견된다. 성체 체세포 조직에서 DNA 메틸화는 일반적으로 CpG 부위(시토신-포스포디에스터 결합-구아닌)에서 발생하며, 비CpG 메틸화는 배아줄기세포(Embryonic stem cell)에서 널리 퍼져 있다.[138][139][140]
포유류에서 모든 CpG 부위의 60~90%는 메틸화되어 있다.[141][142] 메틸화된 시토신 잔기는 자발적으로 아미노기가 제거되어 티민 잔기가 되는데, CpG 이중 핵산염기는 TpG 이중 핵산염기로 변이된다. 이는 인간 게놈에서 CpG 이중 핵산염기의 출현 빈도가 낮다는 점에서 명확하다(CpG 이중 핵산염기는 예상 빈도의 21%만 존재한다).[143] 반면, 비메틸화 시토신의 자발적인 탈아미노화에서는 우라실 잔기가 생성되지만, 이 변이는 세포에 의해 빠르게 인식되어 복구된다.
비메틸화 CpG는 종종 많은 유전자의 5' 조절 영역에 존재하는 '''CpG 섬'''이라고 불리는 클러스터로 모여 있다. 암 등 많은 질병 과정에서 유전자 프로모터인 CpG 섬이 비정상적인 과메틸화를 받아 결과적으로 세포 분열에 의한 자세포에 전달되는 유전자 사일렌싱이 일어난다. DNA 메틸화의 변화는 암 발생의 중요한 요소로 인식되고 있다. 과메틸화가 프로모터와 관련되어 종양 억제 유전자의 사일렌싱을 일으키는 반면, 저메틸화는 일반적으로 초기 발생하는 염색체 불안정성이나 각인 상실과 관련이 있다. 그러나 저메틸화는 후성유전학적 치료의 표적이 될 수도 있다.[144]
DNA 메틸화는 두 가지 방법으로 유전자 전사에 영향을 미친다. 하나는 DNA의 메틸화 자체가 물리적으로 전사 단백질의 유전자 결합을 방해하는 것이고, 다른 하나는 메틸화 DNA가 메틸화 CpG 결합 도메인 단백질(MBD)과 결합하는 것이다. MBD 단백질은 히스톤을 변형하는 히스톤 탈아세틸화 효소 및 기타 크로마틴 재구성 단백질 등 추가적인 단백질을 모집하여 불활성화된 크로마틴(사일런트 크로마틴)을 형성한다. 특히, MeCP2(MeCP2)의 결손은 레트 증후군과 관련이 있으며, MBD2(MBD2)는 암에서 과메틸화 유전자의 전사 사일렌싱을 매개한다.
지금까지의 연구에 따르면, 인간의 장기 기억 유지가 DNA 메틸화에 의해 조절된다는 것이 시사되고 있다.[145][146]
DNA 메틸화는 유전자 발현 조절, 세포 분화, 발생, 질병 등 다양한 생명 현상에 관여하는 중요한 후성유전학적 기전이다. DNA 메틸화 연구는 암을 비롯한 여러 질병의 진단 및 치료 기술 개발에 기여할 수 있을 것으로 기대된다.
한국의 전망: 한국에서는 DNA 메틸화 연구가 활발히 진행되고 있으며, 특히 암 조기 진단 및 치료 기술 개발 분야에서 괄목할 만한 성과가 기대된다.
[1]
논문
The occurrence of 6-methylaminopurine in deoxyribonucleic acids
//www.ncbi.nlm.nih.g[...]
1958-04-01
[2]
논문
Rare bases in animal DNA
1970-01-01
[3]
논문
DNA methylation in thermophilic bacteria: N6-methylcytosine, 5-methylcytosine, and N6-methyladenine.
//www.ncbi.nlm.nih.g[...]
1985-01-01
[4]
논문
Methylation of the deoxyribonucleic acid of Physarum polycephalum at various periods during the mitotic cycle
1970-12-01
[5]
논문
Lack of 5-methylcytosine in Dictyostelium discoideum DNA
1991-07-01
[6]
논문
The ''Dictyostelium discoideum'' genome lacks significant DNA methylation and uncovers palindromic sequences as a source of false positives in bisulfite sequencing
2023-04-01
[7]
논문
Trace analysis of methylated and hydroxymethylated cytosines in DNA by isotope-dilution LC-MS/MS: first evidence of DNA methylation in Caenorhabditis elegans
2015-01-01
[8]
논문
Molecular biology. Methylation talk between histones and DNA
2001-12-01
[9]
논문
Cytosine DNA methylation is found in Drosophila melanogaster but absent in Saccharomyces cerevisiae, Schizosaccharomyces pombe, and other yeast species
2014-04-01
[10]
논문
N6-methyladenine: the other methylated base of DNA
2006-03-01
[11]
논문
DNA methylation on N(6)-adenine in mammalian embryonic stem cells
2016-04-01
[12]
웹사이트
Uracil in DNA: error or signal?
http://www.sciencein[...]
[13]
논문
Reviving the RNA World: An Insight into the Appearance of RNA Methyltransferases
2016-01-01
[14]
논문
De novo methylation of MMLV provirus in embryonic stem cells: CpG versus non-CpG methylation
2002-05-01
[15]
논문
Allele-specific non-CpG methylation of the Nf1 gene during early mouse development
2001-12-01
[16]
논문
Human DNA methylomes at base resolution show widespread epigenomic differences
2009-11-01
[17]
논문
Global epigenomic reconfiguration during mammalian brain development
2013-08-01
[18]
논문
Whole-genome fingerprint of the DNA methylome during human B cell differentiation
2015-07-01
[19]
논문
DNA methylation: an introduction to the biology and the disease-associated changes of a promising biomarker
2010-01-01
[20]
논문
DNA-binding factors shape the mouse methylome at distal regulatory regions
2011-12-01
[21]
논문
Genome-wide evolutionary analysis of eukaryotic DNA methylation
2010-05-01
[22]
논문
CpG methylation is targeted to transcription units in an invertebrate genome
2007-05-01
[23]
논문
Initial sequencing and analysis of the human genome
2001-02-01
[24]
논문
CpG-rich islands and the function of DNA methylation
1986-05-15
[25]
논문
CpG islands in vertebrate genomes
1987-07-01
[26]
논문
Orphan CpG islands identify numerous conserved promoters in the mammalian genome
2010-09-01
[27]
논문
A genome-wide analysis of CpG dinucleotides in the human genome distinguishes two distinct classes of promoters
2006-01-01
[28]
논문
Conservation and divergence of methylation patterning in plants and animals
2010-05-01
[29]
논문
Lineage-specific polycomb targets and de novo DNA methylation define restriction and potential of neuronal progenitors
2008-06-01
[30]
논문
Distribution, silencing potential and evolutionary impact of promoter DNA methylation in the human genome
2007-04-01
[31]
논문
Function and information content of DNA methylation
2015-01-01
[32]
논문
Genome-wide conserved consensus transcription factor binding motifs are hyper-methylated
2010-09-01
[33]
논문
Genome-wide analysis in the mouse embryo reveals the importance of DNA methylation for transcription integrity
2020-06-01
[34]
논문
Dnmt1-independent CG methylation contributes to nucleosome positioning in diverse eukaryotes
2014-03-01
[35]
논문
Cytosine methylation and the ecology of intragenomic parasites
1997-08-01
[36]
논문
DNA methylation enables transposable element-driven genome expansion
2020-08-00
[37]
논문
The alternative role of DNA methylation in splicing regulation
2015-05-00
[38]
논문
Conserved role of intragenic DNA methylation in regulating alternative promoters
2010-07-00
[39]
논문
Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription
2005-11-00
[40]
논문
Programming of DNA methylation patterns
https://zenodo.org/r[...]
2012-07-00
[41]
논문
Global DNA methylation and chondrogenesis of rat limb buds in a three-dimensional organ culture system
2022-07-00
[42]
논문
Loss of methylation activates Xist in somatic but not in embryonic cells
1995-10-00
[43]
논문
Role for DNA methylation in genomic imprinting
1993-11-00
[44]
논문
Targets and dynamics of promoter DNA methylation during early mouse development
2010-12-00
[45]
논문
Reprogramming DNA methylation in the mammalian life cycle: building and breaking epigenetic barriers
2013-01-00
[46]
논문
Metabolic recoding of epigenetics in cancer
2018-05-00
[47]
논문
Specific gene hypomethylation and cancer: new insights into coding region feature trends
2009-04-00
[48]
서적
Epigenetics: A Reference Manual
Caister Academic Press
[49]
논문
Epigenetic alterations in aging
2010-08-00
[50]
논문
DNA methylation polymorphisms precede any histological sign of atherosclerosis in mice lacking apolipoprotein E
2004-07-00
[51]
논문
Increased homocysteine and S-adenosylhomocysteine concentrations and DNA hypomethylation in vascular disease
2003-08-00
[52]
논문
Hypermethylation of estrogen receptor-alpha gene in atheromatosis patients and its correlation with homocysteine
2009-10-00
[53]
논문
DNA methylation and atherosclerosis
2002-08-00
[54]
논문
Methylation of the estrogen receptor-alpha gene promoter is selectively increased in proliferating human aortic smooth muscle cells
2000-04-00
[55]
논문
Inactivation of monocarboxylate transporter MCT3 by DNA methylation in atherosclerosis
2005-08-00
[56]
논문
Genome-wide DNA methylation encodes cardiac transcriptional reprogramming in human ischemic heart failure
2019-03-00
[57]
논문
Racial and socioeconomic disparity associates with differences in cardiac DNA methylation among men with end-stage heart failure
2021-05-00
[58]
논문
DNA Methylation Clocks in Aging: Categories, Causes, and Consequences
2018-09-00
[59]
논문
A longitudinal study of epigenetic variation in twins
2010-08-00
[60]
논문
Distinct DNA methylomes of newborns and centenarians
2012-06-00
[61]
웹사이트
The lipid elongation enzyme ELOVL2 is a molecular regulator of aging in the retina
https://onlinelibrar[...]
2020-02-00
[62]
웹사이트
Impact of age, BMI and HbA1c levels on the genome-wide DNA methylation and mRNA expression patterns in human adipose tissue and identification of epigenetic biomarkers in blood
https://academic.oup[...]
2015-04-10
[63]
논문
Acute exercise remodels promoter methylation in human skeletal muscle
2012-03-00
[64]
논문
A six months exercise intervention influences the genome-wide DNA methylation pattern in human adipose tissue
2013-06-00
[65]
논문
Physical activity and global genomic DNA methylation in a cancer-free population
2011-03-00
[66]
논문
Dynamic DNA methylation controls glutamate receptor trafficking and synaptic scaling
2016-05-00
[67]
논문
Epigenetic regulation and chromatin remodeling in learning and memory
2017-01-00
[68]
논문
Memory consolidation for contextual and auditory fear conditioning is dependent on protein synthesis, PKA, and MAP kinase
1999-00-00
[69]
논문
DNA methylation changes in plasticity genes accompany the formation and maintenance of memory
2016-01-00
[70]
논문
Experience-dependent epigenomic reorganization in the hippocampus
2017-07-00
[71]
논문
DNA Methylation and Establishing Memory
2022
[72]
웹사이트
Review on DNA Methylation
http://www.methods.i[...]
[73]
논문
The DNA methyltransferase DNMT3C protects male germ cells from transposon activity
2016-11-00
[74]
논문
rahu is a mutant allele of Dnmt3c, encoding a DNA methyltransferase homolog required for meiosis and transposon repression in the mouse male germline
2017-08-00
[75]
논문
Methylation of tRNAAsp by the DNA methyltransferase homolog Dnmt2
2006-01-00
[76]
논문
Dynamics and function of DNA methylation in plants
2018-08-00
[77]
논문
DNA methylation in Arabidopsis has a genetic basis and shows evidence of local adaptation
2015-05-00
[78]
논문
Locus-specific control of asymmetric and CpNpG methylation by the DRM and CMT3 methyltransferase genes
2002-12-00
[79]
논문
Widespread natural variation of DNA methylation within angiosperms
2016-09-00
[80]
논문
Genome-wide evolutionary analysis of eukaryotic DNA methylation
2010-05-00
[81]
논문
RNA-directed DNA methylation in Arabidopsis
2002-12-00
[82]
논문
Evolution of DNA Methylation across Insects
2017-03-00
[83]
논문
Functional CpG methylation system in a social insect
2006-10-00
[84]
서적
Physiological and Molecular Mechanisms of Nutrition in Honey Bees
[85]
논문
RNA interference knockdown of DNA methyl-transferase 3 affects gene alternative splicing in the honey bee
2013-07-00
[86]
논문
The two-dimensional restriction analysis of Drosophila DNAs: males and females
1981-05-00
[87]
논문
DNA methylation in Drosophila melanogaster
2000-11-00
[88]
논문
Genome methylation in D. melanogaster is found at specific short motifs and is independent of DNMT2 activity
2014-05-00
[89]
논문
N6-methyladenine DNA modification in Drosophila
2015-05-00
[90]
논문
DNA methylation in the fungi
1984-07-00
[91]
논문
A comparison of DNA methylation levels in selected isolates of higher fungi
[92]
논문
Bisulfite sequencing reveals that Aspergillus flavus holds a hollow in DNA methylation
2012-00-00
[93]
논문
The methylated component of the Neurospora crassa genome
2003-04-00
[94]
논문
Lack of 5-methylcytosine in Dictyostelium discoideum DNA
1991-07-00
[95]
논문
Methjylation in Physarum DNA
1980-07-00
[96]
논문
The Epigenomic Landscape of Prokaryotes
2016-02-00
[97]
논문
Bacterial Epigenomics: Coming of Age
2021-08-00
[98]
논문
Epigenomic characterization of Clostridioides difficile finds a conserved DNA methyltransferase that mediates sporulation and pathogenesis
2020-01-00
[99]
논문
The dam and dcm strains of Escherichia coli--a review
1994-05-00
[100]
웹사이트
Making unmethylated (dam-/dcm-) DNA
http://www.neb.com/n[...]
[101]
논문
Crime investigation through DNA methylation analysis: methods and applications in forensics.
2018-01-00
[102]
논문
Optimizing methodologies for PCR-based DNA methylation analysis
2013-10-00
[103]
논문
Efficient and accurate determination of genome-wide DNA methylation patterns in Arabidopsis thaliana with enzymatic methyl sequencing
2020-10-07
[104]
논문
Kinetic analysis of Yersinia pestis DNA adenine methyltransferase activity using a hemimethylated molecular break light oligonucleotide
2007-08-00
[105]
논문
Hairpin fluorescence DNA probe for real-time monitoring of DNA methylation
2007-02-00
[106]
논문
Methylation-sensitive high resolution melting (MS-HRM): a new approach for sensitive and high-throughput assessment of methylation
[107]
논문
Quantitative evaluation of DNA methylation by optimization of a differential-high resolution melt analysis protocol
2009-07-00
[108]
논문
Reconstructing the DNA methylation maps of the Neandertal and the Denisovan
2014-05-00
[109]
논문
Reconstructing Denisovan Anatomy Using DNA Methylation Maps
2019-09-00
[110]
논문
Methylation Markers for the Identification of Body Fluids and Tissues from Forensic Trace Evidence
2016-02-01
[111]
웹사이트
Infinium Methylation Assay Interrogate single CpG sites
https://www.illumina[...]
2020-01-10
[112]
웹사이트
Infinium MethylationEPIC Kit Methylation profiling array for EWAS
https://www.illumina[...]
2020-01-10
[113]
논문
An integrated resource for genome-wide identification and analysis of human tissue-specific differentially methylated regions (tDMRs)
2008-09-00
[114]
논문
Potential forensic application of DNA methylation profiling to body fluid identification
2012-01-00
[115]
논문
The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores
2009-02-00
[116]
논문
Epigenetic reprogramming in mammalian development
2001-08-00
[117]
논문
Genome-scale DNA methylation maps of pluripotent and differentiated cells
2008-08-00
[118]
논문
Differential methylation of tissue- and cancer-specific CpG island shores distinguishes human induced pluripotent stem cells, embryonic stem cells and fibroblasts
2009-12-00
[119]
논문
Intra-individual change over time in DNA methylation with familial clustering
2008-06-00
[120]
논문
Inter-individual variation of DNA methylation and its implications for large-scale epigenome mapping
2008-06-00
[121]
웹사이트
QDMR: a quantitative method for identification of differentially methylated regions by entropy
http://bioinfo.hrbmu[...]
2013-03-09
[122]
논문
QDMR: a quantitative method for identification of differentially methylated regions by entropy
2011-05-00
[123]
논문
Gene-set analysis is severely biased when applied to genome-wide methylation data
2013-08-00
[124]
논문
Systematic identification and annotation of human methylation marks based on bisulfite sequencing methylomes reveals distinct roles of cell type-specific hypomethylation in the regulation of cell identity genes
2016-01-00
[125]
웹사이트
SMART 2: A Comprehensive Analysis Tool for Bisulfite Sequencing Data
http://fame.edbc.org[...]
2016-00-00
[126]
논문
Molecular approaches for forensic cell type identification: On mRNA, miRNA, DNA methylation and microbial markers
2015-09-00
[127]
논문
DNA methylation and application in forensic sciences
2015-04-00
[128]
논문
Developmental validation studies of epigenetic DNA methylation markers for the detection of blood, semen and saliva samples
2016-07-00
[129]
논문
Principles of DNA methylation and their implications for biology and medicine
2018-09-00
[130]
논문
Sensitive and specific multi-cancer detection and localization using methylation signatures in cell-free DNA
2020-06-00
[131]
논문
Comprehensive human cell-type methylation atlas reveals origins of circulating cell-free DNA in health and disease
2018-11-00
[132]
논문
Prediction of methylated CpGs in DNA sequences using a support vector machine
https://eprints.ucm.[...]
2005-08-00
[133]
논문
CpG island methylation in human lymphocytes is highly correlated with DNA sequence, repeats, and predicted DNA structure
2006-03-00
[134]
논문
Enhancement on the predictive power of the prediction model for human genomic DNA methylation
[135]
논문
CpGIMethPred: computational model for predicting methylation status of CpG islands in human genome
[136]
논문
Reprogramming of the paternal genome upon fertilization involves genome-wide oxidation of 5-methylcytosine
[137]
논문
Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals
[138]
논문
De novo methylation of MMLV provirus in embryonic stem cells: CpG versus non-CpG methylation
[139]
논문
Allele-Specific Non-CpG Methylation of the Nf1 Gene during Early Mouse Development
[140]
논문
Human DNA methylomes at base resolution show widespread epigenomic differences
[141]
논문
Amount and distribution of 5-methylcytosine in human DNA from different types of tissues or cells
[142]
논문
Methylated cytosine and the brain: a new base for neuroscience
[143]
논문
Initial sequencing and analysis of the human genome
[144]
논문
Specific gene hypomethylation and cancer: New insights into coding region feature trends
[145]
논문
Covalent modification of DNA regulates memory formation
2007-03-15
[146]
웹사이트
Memories may be stored on your DNA
http://www.newscient[...]
New Scientist
2008-12-02
[147]
서적
Epigenetics: A Reference Manual
Caister Academic Press
[148]
서적
がんの分子生物学
メディカル・サイエンス・インターナショナル
[149]
논문
ゲノムDNAのメチル化修飾の形成と維持の機構
http://lifesciencedb[...]
2012-12-05
[150]
논문
Methylation of tRNAAsp by the DNA methyltransferase homolog Dnmt2
[151]
논문
Locus-specific control of asymmetric and CpNpG methylation by the DRM and CMT3 methyltransferase genes
[152]
웹사이트
ChromDB::Chromatin Database
http://chromdb.org
2011-04-05
[153]
논문
RNA-directed DNA methylation in Arabidopsis
[154]
논문
DNA methylation in the fungi
[155]
논문
A comparison of DNA methylation levels in selected isolates of higher fungi
Mycological Society of America
[156]
논문
Stage-specific DNA methylation in a fungal plant pathogen
http://jb.asm.org/cg[...]
[157]
논문
The methylated component of the Neurospora crassa genome
[158]
논문
Diverse DNA modification in marine prokaryotic and viral communities
https://doi.org/10.1[...]
2022-01-21
[159]
논문
The DNA methylation landscape of giant viruses
https://www.nature.c[...]
2020-05-27
[160]
논문
The dam and dcm strains of ''Escherichia coli''—a review
[161]
웹사이트
Making unmethylated (dam-/dcm-) DNA
http://www.neb.com/n[...]
2011-04-05
[162]
논문
Shotgun bisulphite sequencing of the Arabidopsis genome reveals DNA methylation patterning.
[163]
논문
Bismark: a flexible aligner and methylation caller for Bisulfite-Seq applications.
[164]
논문
BSMAP: whole genome bisulfite sequence MAPping program.
[165]
논문
Kinetic analysis of Yersinia pestis DNA adenine methyltransferase activity using a hemimethylated molecular break light oligonucleotide
[166]
논문
Hairpin fluorescence DNA probe for real-time monitoring of DNA methylation
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com